|
Description
Edmontosaurus has been described in detail from several specimens.
Like other hadrosaurids, it was a bulky animal with a long, laterally
flattened tail and a head with an expanded, duck-like beak. The skull had no
hollow or solid crest, unlike many other hadrosaurids. The fore legs were
not as heavily built as the hind legs, but were long enough to be used in
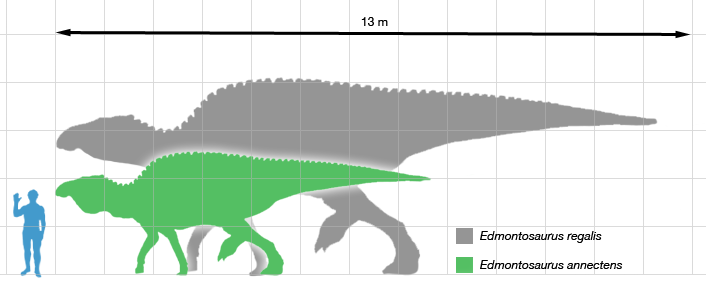
standing or movement. Edmontosaurus was among the largest
hadrosaurids: depending on the species, a fully grown adult could have been
9 meters (30 ft) long, and some of the larger specimens reached the range of
12 meters (39 ft)
to 13 meters (43 ft) long.
Its weight was on the order of 4.0 metric tons (4.4 short tons).
Skull
The skull of a fully grown
Edmontosaurus was around a meter (or
yard) long.
The skull was roughly triangular in profile,
with no bony cranial crest.
Viewed from above, the front and rear of the skull were expanded, with the
broad front forming a duck-bill or
spoon-bill shape. The beak was toothless, and both the upper and lower
beaks were extended by
keratinous
material.
Substantial remains of the keratinous upper beak are known from the
"mummy" kept at the
Senckenberg Museum.
In this specimen, the preserved nonbony part of the beak extended for at
least 8 centimeters (3.1 in) beyond the bone, projecting down vertically.
The nasal openings of
Edmontosaurus were elongate and housed in deep
depressions surrounded by distinct bony rims above, behind, and below.
Teeth were present only in the maxillae (upper cheeks) and dentaries
(main bone of the lower jaw). The teeth were continually replaced, taking
about half a year to form.
They grew in columns, with an observed maximum of six in each, and the
number of columns varied based on the animal's size.
Known column counts for the various species are: 51 to 53 columns per
maxilla and 48 to 49 per dentary (teeth of the upper jaw being slightly
narrower than those in the lower jaw) for E. regalis; 43 columns per
maxilla and 36 per dentary for E. annectens; and 52 and 44 for
E.
saskatchewanensis.
Skin
Multiple specimens of
Edmontosaurus have been found with preserved
skin impressions. Several have been well-publicized, such as the "Trachodon
mummy" of the early 20th century,
and the specimen nicknamed "Dakota",
the latter apparently including remnant
organic compounds from the skin.
Because of these finds, the
scalation of
Edmontosaurus is known for most areas of the body.
AMNH 5060, the "Trachodon mummy" (so-called because it appears to be a
fossil of a
natural mummy), is now recognized as a specimen of
E. annectens.
It was found to have skin impressions over the snout, much of the neck and
torso, and parts of the arms and legs.
The tail and part of the legs eroded before collection, so these areas are
unknown for the specimen.
Additionally, some areas with skin impressions, such as sections associated
with the neck ridge (see below) and hands, were accidentally removed during
preparation of the specimen.
The specimen is thought to have desiccated in a dry stream bed,
probably on or near a
point bar.
The circumstances of the location and preservation of the body suggest that
the animal died during a prolonged drought, perhaps from starvation.
The desiccated carcass was eventually buried in a sudden flood, surrounded
by sediment that had enough
fine particles
to make a cast of the epidermal structures.
The
epidermis was thin, and the scalation composed of small nonoverlapping
scales,
as seen in the
Gila monster.
Two general types of scales were present over most of the body: small
pointed or convex
tubercles,
1 to 3 millimeters (0.039 to 0.12 in) in diameter with no definite
arrangement (ground tubercles); and larger, flat
polygonal
tubercles (pavement tubercles) typically less than 5 millimeters (0.20 in)
in diameter, but up to 10 millimeters (0.39 in) over the forearm. The
pavement tubercles were grouped into clusters separated by ground tubercles,
with transitional scales between the two types. Over most of the body, the
pavement tubercles were arranged in circular or oval clusters, while near
the shoulder on the upper arm, they formed strips roughly parallel to each
other and the shoulder blade. Generally, clusters were larger on the upper
surfaces of the body and smaller on the underside. Clusters up to 50
centimeters (20 in) in length were present above the hips.
Paleobiology
Brain and nervous
system
The brain of
Edmontosaurus has been described in several papers
and abstracts through
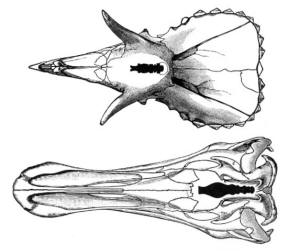 |
| A 1905 chart showing the relatively small brains of a
Triceratops (top) and Edmontosaurus |
the use of
endocasts of the cavity where the brain had been.
E. annectens
and E. regalis,
as well as specimens not identified to species,
have been studied in this way. The brain was not particularly large for an
animal the size of Edmontosaurus. The space holding it was only about
a quarter of the length of the skull,
and various endocasts have been measured as displacing 374 milliliters (13
US fl oz)
to 450 milliliters (15 US fl oz),
which does not take into account that the brain may have occupied as little
as 50% of the space of the endocast, the rest of the space being taken up by
the
dura mater surrounding the brain.
Diet
Feeding adaptations
As a hadrosaurid,
Edmontosaurus was a large terrestrial
herbivore. Its teeth were continually replaced and packed into dental
batteries that contained hundreds of teeth, only a relative handful of which
were in use at any time.
It used its broad beak to cut loose food, perhaps by cropping,
or by closing the jaws in a clamshell-like manner over twigs and branches
and then stripping off the more nutritious leaves and shoots.
Because the tooth rows are deeply indented from the outside of the jaws, and
because of other anatomical details, it is inferred that
Edmontosaurus
and most other ornithischians had cheek-like structures, muscular or
non-muscular. The function of the cheeks was to retain food in the mouth.
The animal's feeding range would have been from ground level to around 4
meters (13 ft) above.
Isotopic studies
The diet and
physiology of
Edmontosaurus have been probed by using
stable isotopes of
carbon and
oxygen as
recorded in
tooth enamel. When feeding, drinking, and breathing, animals take in
carbon and oxygen, which become incorporated into bone. The isotopes of
these two elements are determined by various internal and external factors,
such as the type of plants being eaten, the physiology of the animal,
salinity,
and climate. If isotope ratios in fossils are not altered by fossilization
and later
changes,
they can be studied for information about the original factors;
warmblooded animals will have certain isotopic compositions compared to
their surroundings, animals that eat certain types of plants or use certain
digestive processes will have distinct isotopic compositions, and so on.
Enamel is typically used because the structure of the mineral that forms
enamel makes it the most resistant material to chemical change in the
skeleton.
A 2004 study by Kathryn Thomas and Sandra Carlson used teeth from the
upper jaw of three individuals interpreted as a juvenile, a subadult, and an
adult, recovered from a bone bed in the Hell Creek Formation of
Corson County, South Dakota. In this study, successive teeth in columns
in the edmontosaurs' dental batteries were sampled from multiple locations
along each tooth using a microdrilling system. This sampling method takes
advantage of the organization of hadrosaurid dental batteries to find
variation in tooth isotopes over a period of time. From their work, it
appears that edmontosaur teeth took less than about 0.65 years to form,
slightly faster in younger edmontosaurs. The teeth of all three individuals
appeared to show variation in oxygen isotope ratios that could correspond to
warm/dry and cool/wet periods; Thomas and Carlson considered the possibility
that the animals were migrating instead, but favored local seasonal
variations because migration would have more likely led to ratio
homogenization, as many animals migrate to stay within specific temperature
ranges or near particular food sources.
Pathologies and
health
In 2003, evidence of
tumors was described in
Edmontosaurus bones.
Researchers tested dinosaur vertebrae for tumors using
computerized tomography and
fluoroscope screening. Several other hadrosaurids, including
Brachylophosaurus,
Gilmoreosaurus, and
Bactrosaurus, also tested positive. Although more than 10,000
fossils were examined in this manner, the tumors were limited to
Edmontosaurus and closely related genera. The tumors may have been
caused by environmental factors or
genetic
propensity.
Osteochondrosis, or surficial pits in bone at places where bones
articulate, is also known in Edmontosaurus. This condition, resulting
from
cartilage failing to be replaced by bone during growth, was found to be
present in 2.2% of 224 edmontosaur toe bones. The underlying cause of the
condition is unknown. Genetic predisposition, trauma, feeding intensity,
alterations in blood supply, excess
thyroid
hormones,
and deficiencies in various growth factors have been suggested. Among
dinosaurs, osteochondrosis (like tumors) is most commonly found in
hadrosaurids.
Locomotion
Like other hadrosaurids,
Edmontosaurus is thought to have been a
facultative biped, meaning that it mostly moved on four legs, but could
adopt a bipedal stance when needed. It probably went on all fours when
standing still or moving slowly, and switched to using the hind legs alone
when moving more rapidly.
Research conducted by computer modeling in 2007 suggests that
Edmontosaurus could run at high speeds, perhaps up to 45 kilometers per
hour (28 mph).
Further simulations using a subadult specimen estimated as weighing 715
kilograms (1,580 lb) when alive produced a model that could run or hop
bipedally, use a
trot,
pace, or single foot symmetric quadrupedal gait, or move at a
gallop.
Interactions
with theropods
The time span and geographic range of
Edmontosaurus overlapped
with
Tyrannosaurus,
 |
| The damage to the tail vertebrae of this Edmontosaurus
skeleton (on display at the Denver Museum of Nature and Science)
indicates that it may have been bitten by a
Tyrannosaurus. (Picture
Source) |
and an adult specimen of
E. annectens on
display in the
Denver Museum of Nature and Science shows evidence of a theropod bite in
the tail. Counting back from the hip, the thirteenth to seventeenth
vertebrae have damaged
spines consistent with an attack from the right rear of the animal. One
spine has a portion sheared away, and the others are kinked; three have
apparent tooth puncture marks. The top of the tail was at least 2.9 meters
(9.5 ft) high, and the only theropod species known from the same rock
formation that was tall enough to make such an attack is
T. rex. The
bones are partially healed, but the edmontosaur died before the traces of
damage were completely obliterated. The damage also shows signs of bone
infection. Kenneth Carpenter, who studied the specimen, noted that there
also seems to be a healed fracture in the left hip which predated the attack
because it was more fully healed. He suggested that the edmontosaur was a
target because it may have been limping from this earlier injury. Because it
survived the attack, Carpenter suggested that it may have outmaneuvered or
outran its attacker, or that the damage to its tail was incurred by the
hadrosaurid using it as a weapon against the tyrannosaur.
Another specimen of
E. annectens, pertaining to a 7.6 meters (25
ft) long individual from South Dakota, shows evidence of tooth marks from
small theropods on its lower jaws. Some of the marks are partially healed.
Michael Triebold, informally reporting on the specimen, suggested a scenario
where small theropods attacked the throat of the edmontosaur; the animal
survived the initial attack but succumbed to its injuries shortly
thereafter.
Some edmontosaur bone beds were sites of scavenging.
Albertosaurus
and
Saurornitholestes tooth marks are common at one Alberta bone bed,
and
Daspletosaurus fed on
Edmontosaurus and fellow hadrosaurid
Saurolophus at another Alberta site.
Social behavior
Extensive
bone beds
are known for Edmontosaurus, and such groupings of hadrosaurids are
used to suggest that they were gregarious, living in groups.
Four quarries containing edmontosaur remains are identified in a 2007
database of fossil bone beds, from Alaska (Prince Creek Formation), Alberta
(Horseshoe Canyon Formation), South Dakota (Hell Creek Formation), and
Wyoming (Lance Formation).
One edmontosaur bone bed, from
claystone
and
mudstone of the Lance Formation in eastern Wyoming, covers more than a
square kilometer, although Edmontosaurus bones are most concentrated
in a 40 hectares (0.15 sq mi) subsection of this site. It is estimated that
disassociated remains pertaining to 10,000 to 25,000 edmontosaurs are
present here.
Because of its wide distribution, which covers a distance from Alaska to
Colorado and includes
polar settings that would have had little light during a significant
part of the year, Edmontosaurus has been considered possibly
migratory. A 2008 review of dinosaur migration studies by Phil R. Bell and
Eric Snively proposed that E. regalis was capable of an annual 2,600
kilometers (1,600 mi) round-trip journey, provided it had the requisite
metabolism and fat deposition rates. Such a trip would have required
speeds of about 2 to 10 kilometers per hour (1 to 6 mph), and could have
brought it from Alaska to Alberta. The possible migratory nature of
Edmontosaurus contrasts with many other dinosaurs, such as
theropods,
sauropods,
and
ankylosaurians, which Bell and Snively found were more likely to have
overwinter.
Return to the
Old Earth Ministries Online Dinosaur
Curriculum homepage.

Shopping
Bay
State Replicas - Skull, Claw
Black
Hills Institute - Skeletons, skull, half-skull, arm, hand, leg, femur,
foot, temenal phalange, juvenile skeleton, juvenile skull
|