|
It can be
distinguished from other abelisaurids by its wider skull, the very rough
texture and thickened bone on the top of its snout, and the single rounded
horn on the roof of its skull, which was
originally mistaken for the dome of a pachycephalosaur. It
also had more teeth in both upper
and lower jaws than most abelisaurids.
Known from several well-preserved
skulls and abundant skeletal material,
Majungasaurus has recently become one of the
best-studied theropod dinosaurs from the Southern Hemisphere.
It appears to be most closely related to abelisaurids from India rather than
South America or continental
Africa, a fact which
has important
biogeographical
implications. Majungasaurus
was the apex predator in its
ecosystem, mainly
preying on sauropods like
Rapetosaurus, and is
also the only dinosaur for which direct evidence of cannibalism is known.
Description
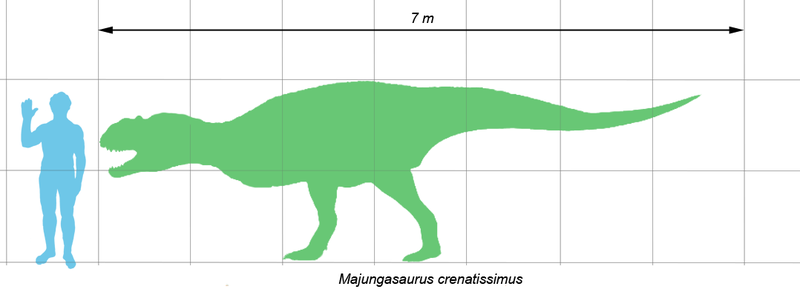
Majungasaurus
was a medium-sized theropod that typically measured 6–7 meters (20–23 ft) in
length, including its tail. Fragmentary remains of larger individuals
indicate that some adults reached lengths of more than 8 meters (26 ft).
Scientists estimate that an average adult
Majungasaurus weighed more than 1100 kilograms
(2400 lb), although the largest animals would have weighed more. Its
8–9 meter (26–30 ft) relative Carnotaurus
has been estimated to weigh 1500 kilograms (3300 lb).
The skull of
Majungasaurus is
exceptionally well-known compared to most theropods and generally similar to
that of other abelisaurids. Like other abelisaurid skulls, its length was
proportionally short for its height, although not as short as in
Carnotaurus. The skulls of
large individuals measured 60–70 centimeters (24–28 in) long. However, the skull of
Majungasaurus was markedly wider than in other
abelisaurids. All abelisaurids had a rough, sculptured texture on the
outside faces of the skull bones, and
Majungasaurus was no exception. This was carried
to an extreme on the nasal bones of
Majungasaurus, which were extremely thick and
fused together, with a low central ridge running along the half of the bone
closest to the nostrils. A distinctive dome-like horn protruded from the
fused
frontal bones on top of
the skull as well. In life, these structures would have been covered with
some sort of
integument, possibly
made of
keratin.
The postcranial skeleton of
Majungasaurus closely
resembles those of Carnotaurus
and Aucasaurus, the
only other abelisaurid genera for which complete skeletal material is known.
Majungasaurus was
bipedal, with a long tail to balance out the head and torso, putting the
center of gravity over the hips. The humerus (upper arm bone)
was short and curved, closely resembling those of
Aucasaurus and
Carnotaurus. Also like
related dinosaurs, Majungasaurus
had very short forelimbs with four extremely reduced digits, with only two
very short external fingers and no claws. The hand and finger bones of
Majungasaurus, like
other carnotaurines, lacked the characteristic pits and grooves where claws
and tendons would normally attach, and its finger bones were fused together,
indicating that the hand was immobile.
Like other abelisaurids, the
hindlimbs were stocky and short compared to body length. The tibia (lower
leg bone) of Majungasaurus
was even stockier than that of its relative
Carnotaurus, with a prominent crest on the knee.
The
astragalus and
calcaneum (ankle bones)
were fused together, and the feet bore three functional digits, with a
smaller first digit that did not contact the ground.
Discovery and Naming
French
paleontologist
Charles Depéret
described the first theropod remains from northwestern Madagascar in 1896.
These included two teeth, a claw, and some vertebrae discovered along the
Betsiboka River by a
French army officer and deposited in the collection of what is now the
Université Claude Bernard Lyon 1. Depéret
referred these fossils to the genus
Megalosaurus, which at the time was a
wastebasket taxon
containing any number of unrelated large theropods, as the new species
M. crenatissimus.
Numerous fragmentary remains from
Mahajanga Province in
northwestern Madagascar were recovered by French collectors over the next
100 years, many of which were deposited in the
Muséum National d'Histoire Naturelle
in Paris. In 1955,
René Lavocat described
a theropod dentary with teeth from the
Maevarano Formation in
the same region where the original material was found. The teeth matched
those first described by Depéret, but the strongly curved jaw bone was very
different from both Megalosaurus
and Dryptosaurus.
Lavocat renamed the genus Majungasaurus,
using an older spelling of Mahajanga as well as the
Greek word meaning
"lizard", and made this jaw bone the type specimen. In the 1990s,
more specimins were found which helped describe Majungasaurus to a greater
extent.
Paleobiology
Skull
ornamentation
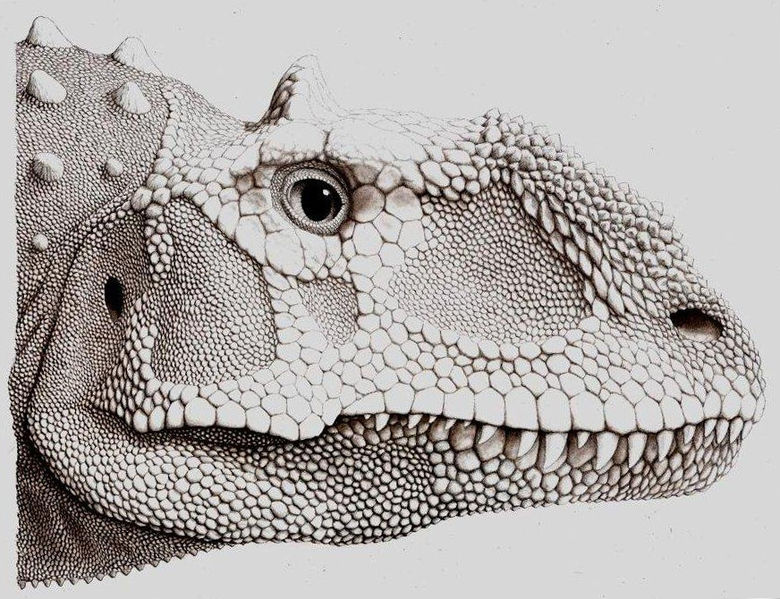
Majungasaurus
is perhaps most distinctive for its skull ornamentation, including the
swollen and fused nasals and the frontal horn. Other ceratosaurs, including
Carnotaurus,
Rajasaurus, and
Ceratosaurus itself
bore crests on the head. These structures are likely to have played a role
in
intraspecific competition,
although their exact function within that context is unknown. The hollow
cavity inside the frontal horn of Majungasaurus
would have weakened the structure and probably precluded its use in direct
physical combat, although the horn may have served a
display purpose.
While there is variation in the ornamentation of
Majungasaurus individuals,
there is no evidence for
sexual dimorphism.
Feeding
Scientists have suggested that the
unique skull shape of Majungasaurus
and other abelisaurids indicate different predatory habits than other
theropods. Whereas most theropods were characterized by long, low skulls of
narrow width, abelisaurid skulls were taller and wider, and often shorter in
length as well. The narrow skulls of other
theropods were well-equipped to withstand the vertical stress of a powerful
bite, but not as good at withstanding torsion (twisting).
In comparison to modern mammalian predators, most theropods may have used a
strategy similar in some ways to that of long- and narrow-snouted
canids, with the
delivery of many bites weakening the prey animal.
|

|
|
Restoration of the head of
Majungasaurus crenatissimus (From
Wikipedia)
|
Abelisaurids, especially
Majungasaurus, may instead
have been adapted for a feeding strategy more similar to modern
felids, with short and
broad snouts, that bite once and hold on until the prey is subdued.
Majungasaurus had an even
broader snout than other abelisaurids, and other aspects of its anatomy may
also support the bite-and-hold hypothesis. The neck was strengthened, with
robust vertebrae, interlocking ribs and ossified tendons, as well as
reinforced muscle attachment sites on the vertebrae and the back of the
skull. These muscles would have been able to hold the head steady despite
the struggles of its prey. Abelisaurid skulls were also strengthened in many
areas by bone
mineralized out of the
skin, creating the
characteristic rough texture of the bones. This is particularly true of
Majungasaurus,
where the nasal bones were fused and thickened for strength. On the other
hand, the lower jaw of Majungasaurus
sported a large
fenestra (opening) on
each side, as seen in other ceratosaurs, as well as
synovial joints between
certain bones that allowed a high degree of flexibility in the lower jaw,
although not to the extent seen in snakes. This may have been an adaptation
to prevent the fracture of the lower jaw when holding onto a struggling prey
animal. The front teeth of the upper jaw were more robust than the rest, to
provide an anchor point for the bite, while the low crown height of
Majungasaurus teeth
prevented them from breaking off during a struggle. Finally, unlike the
teeth of Allosaurus
and most other theropods, which were curved on both the front and back,
abelisaurids like Majungasaurus
had teeth curved on the front edge but straighter on the back (cutting)
edge. This structure may have served to prevent slicing, and instead holding
the teeth in place when biting.
Majungasaurus
was the largest predator in its environment, while the only known large
herbivores at the time were sauropods like
Rapetosaurus. Scientists have suggested that
Majungasaurus, and
perhaps other abelisaurids, specialized on hunting sauropods.
Majungasaurus
tooth marks on Rapetosaurus
bones confirm that it at least fed on these sauropods, whether or not it
actually killed them.
Cannibalism
Although sauropods may have been the
prey of choice for Majungasaurus,
recent discoveries in Madagascar indicate another surprising component of
its diet: other Majungasaurus.
Numerous bones of Majungasaurus
have been discovered bearing tooth marks identical to those found on
sauropod bones from the same localities. These marks have the same spacing
as teeth in Majungasaurus
jaws, are of the same size as Majungasaurus
teeth, and contain smaller notches consistent with the serrations on those
teeth. As Majungasaurus
is the only large theropod known from the area, the simplest explanation is
that it was feeding on other members of its own species.
Suggestions that the Triassic
Coelophysis was a
cannibal have been recently disproven, leaving
Majungasaurus as the only non-avian theropod
with confirmed cannibalistic tendencies,
although there is some evidence that cannibalism may have occurred in other
species as well.
It is unknown if
Majungasaurus actively
hunted their own kind or only scavenged their carcasses.
Respiratory
System
|
|
|
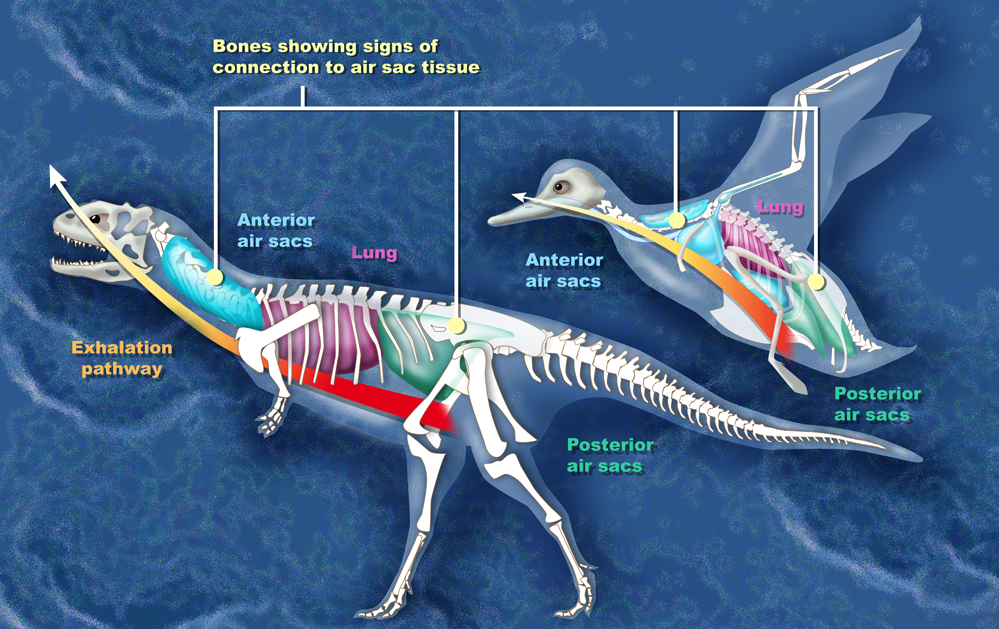
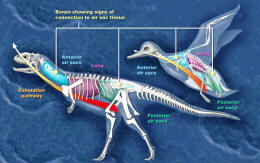
Comparison between the air sacs of
Majungasaurus and
a bird
|
Scientists have reconstructed the
respiratory system of Majungasaurus
based on a superbly preserved series of vertebrae (UA 8678) recovered from
the Maevarano Formation. Most of these vertebrae and some of the ribs
contained cavities (pneumatic
foramina) that may have
resulted from the infiltration of
avian-style
lungs and air sacs. In
birds, the neck vertebrae and ribs are hollowed out by the cervical air sac,
the upper back vertebrae by the lung, and the lower back and
sacral (hip) vertebrae
by the abdominal air sac. Similar features in
Majungasaurus vertebrae imply the presence of
these air sacs. These air sacs may have allowed for a basic form of
avian-style 'flow-through ventilation,' where air flow through the lungs is
one-way, so that oxygen-rich air inhaled from outside the body is never
mixed with exhaled air laden with carbon dioxide. This method of
respiration, while complicated, is highly efficient.
The recognition of pneumatic
foramina in Majungasaurus,
besides providing an understanding of its respiratory biology, also has
larger-scale implications for evolutionary biology. The split between the
ceratosaur line, which led to Majungasaurus,
and the
tetanuran line, to
which birds belong, occurred very early in the history of theropods. The
avian respiratory system, present in both lines, must therefore have evolved
before the split, and well before the evolution of birds themselves. This
provides further evidence of the dinosaurian
origin of birds.
Brain and inner ear structure
Computed tomography, also known as
CT scanning, of a complete Majungasaurus
skull (FMNH PR 2100) allowed a rough reconstruction of its brain and inner
ear structure. Overall, the brain was very small relative to body size, but
otherwise similar to many other non-coelurosaurian
theropods, with a very conservative form closer to modern crocodilians than
to birds. One difference between Majungasaurus
and other theropods was its smaller
flocculus, a region of
the
cerebellum that helps
to coordinate movements of the eye with movements of the head. This suggests
that Majungasaurus
and other abelisaurids like
Indosaurus, which also
had a small flocculus, did not rely on quick head movements to sight and
capture prey.
Inferences about behavior can also
be drawn from examination of the inner ear. The
semicircular canals
within the inner ear aid in balance, and the
lateral semicircular canal
is usually parallel to the ground when the animal holds its head in an alert
posture. When the skull of Majungasaurus
is rotated so that its lateral canal is parallel to the ground, the entire
skull is nearly horizontal. This contrasts with many other theropods, where
the head was more strongly downturned when in the alert position. The
lateral canal is also significantly longer in
Majungasaurus than in its more
basal relative
Ceratosaurus,
indicating a greater sensitivity to side-to-side motions of the head.
Pathology
A 2007 report described
pathologies in the
bones of Majungasaurus.
Scientists examined the remains of at least 21 individuals and discovered
four with noticeable pathologies. While
pathology had been studied in large
tetanuran theropods
like
allosaurids and
tyrannosaurids, this
was the first time an abelisauroid had been examined in this manner. No
wounds were found on any skull elements, in contrast to tyrannosaurids where
sometimes gruesome facial bites were common. One of the specimens was a
phalanx (toe bone) of the foot, which had apparently been broken and
subsequently healed.
Most of the pathologies occurred on
the vertebrae. For example, a dorsal (back) vertebra from a juvenile animal
showed an
exostosis (bony growth)
on its underside. The growth probably resulted from the conversion of
cartilage or a
ligament to bone during
development, but the cause of the ossification was not determined.
Hypervitaminosis A and
bone spurs were ruled
out, and an
osteoma (benign bone
tumor) was deemed unlikely. Another specimen, a small caudal (tail)
vertebra, was also found to have an abnormal growth, this time on the top of
its
neural spine, which
projects upwards from the vertebrae, allowing muscle attachment.
Similar growths from the neural spine have been found in specimens of
Allosaurus and
Masiakasaurus, probably
resulting from the ossification of a ligament running either between the
neural spines (interspinal
ligament) or along their tops (supraspinal
ligament).
The most serious pathology
discovered was in a series of five large tail vertebrae. The first two
vertebrae showed only minor abnormalities with the exception of a large
groove that extended along the left side of both bones. However, the next
three vertebrae were completely fused together at many different points,
forming a solid bony mass. There is no sign of any other vertebrae after the
fifth in the series, indicating that the tail ended there prematurely. From
the size of the last vertebrae, scientists judged that about ten vertebrae
were lost. One explanation for this pathology is severe physical trauma
resulting in the loss of the tail tip, followed by
osteomyelitis
(infection) of the last remaining vertebrae. Alternatively, the infection
may have come first and led to the end of the tail becoming
necrotic and falling
off. This is the first example of tail truncation known in a non-avian
theropod dinosaur.
Paleoecology
All specimens of
Majungasaurus have been
recovered from the Maevarano Formation in the
Mahajanga Province in
northwestern Madagascar. Most of these, including all of the most complete
material, came from the Anembalemba Member, although
Majungasaurus teeth have
also been found in the underlying Masorobe Member and the overlying Miadana
Member. While these sediments have not been dated
radiometrically,
evidence from
biostratigraphy and
paleomagnetism suggest
that they were deposited during the
Maastrichtian
stage, which lasted
from 70 to 65 Ma (million years ago).
Majungasaurus teeth are found up until the very
end of the Maastrichtian, when all non-avian dinosaurs
went extinct.
Then as now, Madagascar was an
island, having separated from the Indian subcontinent less than 20 million
years earlier. It was drifting northwards but still 10–15° more southerly in
latitude than it is today. The prevailing climate of the time was semi-arid,
with pronounced seasonality in temperature and rainfall.
Majungasaurus inhabited a
coastal flood plain cut by many sandy river
channels. Strong geological evidence suggests
the occurrence of periodic
debris flows through
these channels at the beginning of the wet season, burying the carcasses of
organisms killed during the preceding dry season and providing for their
exceptional preservation as fossils. Sea levels
in the area were rising throughout the Maastrichtian, and would continue to
do so into the
Paleocene Epoch, so
Majungasaurus may
have roamed coastal environments like
tidal flats as well.
The neighboring
Berivotra Formation
represents the contemporaneous marine environment.
Besides
Majungasaurus, fossil taxa
recovered from the Maevarano include fish, frogs, lizards, snakes,
seven distinct species of
crocodylomorphs, five
or six species of mammals,
Vorona and several
other birds, the possibly flighted
dromaeosaurid
Rahonavis, the
noasaurid
Masiakasaurus and two
titanosaurian sauropods,
including Rapetosaurus.
Majungasaurus was by far
the largest carnivore and probably the dominant predator on land, although
large crocodylomorphs like
Mahajangasuchus and
Trematochampsa might
have competed with it closer to water.
If you ordered the Test Pack, it is now time to
take Test 2.
Return to the
Old Earth Ministries Online Dinosaur
Curriculum homepage.

Shopping
|