|
Discovered in 1834 by Johann
Friedrich Engelhardt and described three years later by
Hermann von Meyer,
Plateosaurus was the fifth named dinosaur genus that is still considered
valid today. It is now among the dinosaurs best known to science,
with over 100 skeletons found, some of them nearly complete. German
palaeontologist
Friedrich August von Quenstedt
nicknamed it the Schwäbischer Lindwurm (Swabian dragon) because it was so
common a fossil in southwestern Germany.
Plateosaurus was a
bipedal
herbivore with a small
skull on a long, mobile neck, sharp but plump plant-crushing teeth, powerful
hind limbs, short but muscular arms and grasping hands with large claws on
three fingers, possibly used for defense and feeding. Unusually for a
dinosaur, Plateosaurus showed strong developmental plasticity: instead of
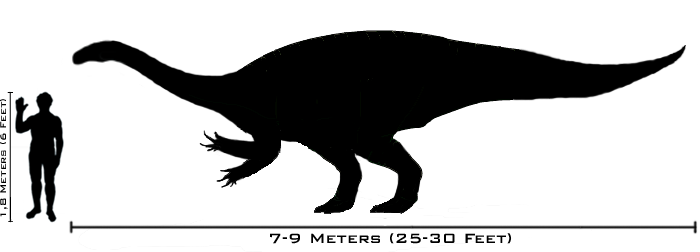
having a fairly uniform adult size, fully grown individuals were between 4.8
and 10 metres (16 and 33 ft) long and weighed between 600 and 4,000
kilograms (1,300 and 8,800 lb). Commonly, the animals lived for 12 to 20
years at least.
Description
Plateosaurus is a member of a group
of early herbivores known as "prosauropods".
The group name is obsolete, as "Prosauropoda" is not a
monophyletic group (thus given in quotation
marks), and most researchers prefer the term basal
sauropodomorph. Plateosaurus had the typical
body shape of a herbivorous bipedal dinosaur: a small skull, a long and
flexible neck composed of ten cervical
(neck) vertebrae, a stocky
body and a long, mobile tail composed of at least 40 caudal
(tail) vertebrae. The arms of Plateosaurus were
very short, even compared to most other "prosauropods", but strongly built,
with hands adapted to powerful grasping. The
shoulder girdle was
narrow (often misaligned in skeletal mounts and drawings),
with the
clavicles (collar
bones) touching at the body midline, as is the
case in other basal sauropodomorphs. The
hindlimbs were held under the body, with slightly flexed knees and ankles,
and the foot
digitigrade – the animal walked on its toes.
The proportionally long lower leg and
metatarsus show that Plateosaurus was
adapted to rapid bipedal locomotion. The tail of
Plateosaurus was typically dinosaurian, muscular and with high mobility.
The skull of Plateosaurus is small
and narrow, rectangular in side view, and nearly three times as long as it
is high. There is an almost rectangular lateral
temporal foramen at the
back, and a large, round
orbit (eye socket), a sub-triangular
antorbital fenestra and
an egg-shaped
naris (nostril) of
almost equal size. The snout carried many small,
leaf-shaped, socketed teeth in both the upper and lower jaw, 5 to 6 on the
premaxilla, 24 to 30 on the
maxilla,
and 21 to 28 on the
dentary. The teeth had
bluntly serrated, thick, leaf-shaped crowns suitable for crushing plant
material. The low position of the jaw joint gave
the chewing muscles great leverage, so that Plateosaurus could deliver a
powerful bite. These features suggest that it
fed exclusively or mainly on plants. Its eyes
were directed to the sides, rather than the front, providing all-round
vision to watch for predators. Some fossil
skeletons have preserved
sclerotic rings.
The ribs were connected to the
dorsal (trunk) vertebrae with two joints, together acting as a simple hinge
joint, which allowed reconstructing the inhaled and exhaled positions of the
ribcage. The difference in volume between both positions determines the air
exchange volume (the amount of air moved with each breath), determined to be
~20 l for a P. engelhardti individual estimated to have weighed 690 kg, or
29 ml/kg bodyweight. This is a typical value for
birds, but not for mammals, and indicates that
Plateosaurus probably had an
avian-style
flow-through lung, although no indicators for
postcranial pneumaticity
(air sacs of the lung invading the bones to reduce weight) can be found on
the bones. Combined with evidence from bone
histology this indicates that Plateosaurus was
an
endotherm.
The
type species of Plateosaurus is
P. engelhardti. Adult individuals of this
species reached 4.8 to 10 metres (16 to 33 ft) in length.
Average individuals had a mass of around 600 to 4,000 kilograms (1,300 to
8,800 lb). The older species, P. gracilis
(formerly named Sellosaurus gracilis), was somewhat smaller, with a total
length of 4 to 5 metres (13 to 16 ft).
Discovery and history
In 1834,
physician Johann Friedrich Engelhardt
discovered some vertebrae and leg bones at
 |
| P. engelhardti, collection number MSF 23 of the Sauriermuseum
Frick, Switzerland, in dorsal view. This is the most complete
Plateosaurus skeleton from Frick. (Picture
Source) |
Heroldsberg near
Nuremberg,
Germany.
Three years later German
palaeontologist
Hermann von Meyer
designated them as the
type
specimen of a new genus, Plateosaurus.
Since then, remains of well over 100 individuals of Plateosaurus have been
discovered at various locations throughout Europe.
Material assigned to Plateosaurus
has been found at over 50 localities in Germany (mainly along the
Neckar
and
Pegnitz river valleys),
Switzerland (Frick) and France.
In 1997, workers of an oil
platform of the
Snorre oilfield located
at the northern end of the
North
Sea were drilling through
sandstone for oil exploration when in a
drill core extracted from 2,256 meters below the seafloor they stumbled upon
a fossil they believed to be plant material. In
2003, the specimen was sent to Jørn Harald Hurum, palaeontologist at the
University of Oslo for
study. Martin Sander and Nicole Klein, palaeontologists of the
University of Bonn,
analyzed the bone microstructure and concluded that the rock preserved
fibrous bone tissue from a fragment of a limb bone belonging to
Plateosaurus, making it the first dinosaur found
in Norway. Plateosaurus material has also been found in Greenland.
Taphonomy
The
taphonomy (burial and fossilization process)
of the three main Plateosaurus sites Trossingen, Halberstadt (both in
Germany) and Frick (Switzerland) is unusual in several ways.
All three sites are nearly monospecific assemblages, meaning that they
contain practically only one species, which requires very special
circumstances. However, shed teeth of
theropods have been found at all three
sites, as well as remains of
Proganochelys, an early turtle.
Additionally, a partial "prosauropod" skeleton was found in Halberstadt that
does not belong to Plateosaurus, but is preserved in a similar position.
All sites yielded almost complete and partial skeletons of Plateosaurus, as
well as isolated bones. The partial skeletons
tend to include the hind limbs and hips, while parts of the anterior body
and neck are rarely found in isolation. The
animals were all
adults or sub-adults;
no juveniles or hatchlings are known. Complete
skeletons and large skeleton parts that include the hind limbs all rest
right side up, as do the turtles. Also, they are
mostly well articulated, and the hind limbs are three-dimensionally
preserved in a zig-zag posture, with the feet often much deeper in the
sediment than the hips.
Earlier interpretations
In the first published discussion of
the Trossingen Plateosaurus finds, Fraas suggested that only miring in mud
allowed the preservation of the single complete skeleton then known.
Similarly, Jaekel interpreted the Halberstadt finds as animals that waded
too deep into swamps, became mired and drowned.
He interpreted partial remains as having been transported into the deposit
by water, and strongly refuted a catastrophic accumulation.
In contrast, von Huene interpreted the sediment as
aeolian deposits, with
the weakest animals, mostly subadults, succumbing to the harsh conditions in
the desert and sinking into the mud of
ephemeral water holes.
He argued that the completeness of many finds indicated that transport did
not happen, and saw partial individuals and isolated bones as results of
weathering and trampling. Seemann developed a
different scenario, in which Plateosaurus herds congregated on large water
holes, and some herd members got pushed in.
Light animals managed to get free, heavy ones got stuck and died.
A different school of thought
developed almost half a century later, with palaeontologist
David Weishampel
suggesting that the skeletons from the lower layers stemmed from a herd that
died catastrophically in a mudflow, while those in the upper layers
accumulated over time. Weishampel explained the
curious monospecific assemblage by theorizing that Plateosaurus were common
during this period. This theory was erroneously
attributed to Seemann in a popular account of the plateosaurs in the
collection of the Institute and Museum for Geology and Paleontology,
University of
Tübingen, and has since
become the standard explanation on most internet sites and in popular books
on dinosaurs. Rieber proposed a more elaborate
scenario, which included the animals dying of thirst or starvation, and
being concentrated by mudflows.
Current interpretation
A detailed re-assessment of the
taphonomy by palaeontologist Martin Sander of the
University of Bonn,
Germany, found that the mud-miring
hypothesis first suggested by Fraas is true:
animals above a certain body weight sank into the mud, which was further
liquefied by their attempts to free themselves. Sander's scenario, similar
to that proposed for the famous
Rancho La Brea Tar Pits,
is the only one explaining all taphonomic data. The degree of completeness
of the carcasses was not influenced by transport, which is obvious from the
lack of indications for transport before burial, but rather by how much the
dead animals were scavenged. Juveniles of Plateosaurus and other taxa of
herbivores were too light to sink into the mud, or managed to extract
themselves, and were thus not preserved. Similarly, the scavenging
theropods were not trapped due to their
lower body weights, combined with a proportionally larger footprint. There
is no indication of herding, nor of catastrophic burial of such a herd, or
catastrophic accumulation of animals that previously died isolated
elsewhere.
Palaeobiology
Posture and gait
From 1980 on, a better understanding
of dinosaur biomechanics, and studies by
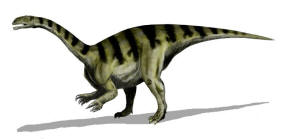 |
| Restoration of P. gracilis, formerly known as Sellosaurus
gracilis (Picture
Source) |
palaeontologists Andreas Christian
and Holger Preuschoft on the resistance to bending of the back of
Plateosaurus, led to widespread acceptance of an
erect, digitigrade limb posture and a roughly horizontal position of the
back. Many researchers were of the opinion that
Plateosaurus could use both quadrupedal gaits (for slow speeds) and bipedal
gaits (for rapid locomotion).
This consensus was changed by a
detailed study of the forelimbs of Plateosaurus by
Bonnan and Senter (2007), which clearly
showed that Plateosaurus was incapable of
pronating its hands.
The pronated position in some museum mounts had been achieved by exchanging
the position of
radius and
ulna
in the elbow. This meant that Plateosaurus was an obligate digitigrade
biped. Further indicators for a purely bipedal mode of locomotion are the
great difference in limb length (the hind limb is roughly twice as long as
the forelimb), the very limited motion range of the forelimb, and the fact
that the
center of mass rests
squarely over the hind limbs.
Feeding and diet
Important cranial characteristics
(such as jaw articulation) of most "prosauropods" are closer to those of
herbivorous reptiles than those of carnivorous ones, and the shape of the
tooth
crown is similar to
those of modern herbivorous or omnivorous
iguanas.
The maximum width of the crown was greater than that of the root, resulting
in a cutting edge similar to those of extant herbivorous or omnivorous
reptiles. This is also true in Plateosaurus.
Paul Barrett proposed that prosauropods supplemented their herbivorous diets
with small prey or
carrion.
So far, no fossil of Plateosaurus
has been found with
gastroliths (gizzard
stones) in the stomach area. The old, widely cited idea that all large
dinosaurs, implicitly also Plateosaurus, swallowed gastroliths to digest
food because of their relatively limited ability to deal with food orally
has been refuted by a study on gastrolith abundance, weight, and surface
structure in fossils compared to alligators and ostriches by Oliver Wings.
The use of gastroliths for digestion seems to have developed on the line
from basal theropods to birds, with a parallel development in
Psittacosaurus.
Growth, metabolism and life span
Similar to all non-avian dinosaurs
studied to date, Plateosaurus grew in a pattern that is dissimilar to both
extant mammals and avian dinosaurs. In the closely related sauropods with
their typical
dinosaurian physiology,
growth was initially rapid, continued somewhat slower well beyond sexual
maturity, but was determinate, i.e. the animals stopped growing at a maximum
size. Mammals grow rapidly, but sexual maturity
falls typically at the end of the rapid growth phase. In both groups, the
final size is relatively constant, with humans atypically variable.
Extant reptiles show a sauropod-like growth
pattern, initially rapid, then slowing after sexual maturity, and almost,
but not fully, stopping in old age. However, their initial growth rate is
much lower than in mammals, birds and dinosaurs. The reptilian growth rate
is also very variable, so that individuals of the same age may have very
different sizes, and final size also varies significantly. In extant
animals, this growth pattern is linked to
behavioural thermoregulation
and a low
metabolic rate (i.e.
ectothermy), and is
called "developmental plasticity". (Note that is
not the same as neural
developmental plasticity.)
Plateosaurus followed a trajectory
similar to sauropods, but with a varied growth rate and final size as seen
in extant reptiles, probably in response to environmental factors such as
food availability. Some individuals were fully grown at only 4.8 metres
(16 ft) total length, while others reached 10 metres (33 ft). However, the
bone microstructure indicates rapid growth, as in sauropods and extant
mammals, which suggests
endothermy. Plateosaurus apparently
represents an early stage in the development of endothermy, in which
endothermy was decoupled from developmental plasticity. This hypothesis is
based on a detailed study of Plateosaurus long-bone histology conducted by
Martin Sander and Nicole Klein of
the University of Bonn,
Germany. A further indication for endothermy is
the avian-style lung of Plateosaurus.
Long-bone histology also allows
estimating the age a specific individual reached. Sander and Klein found
that some individuals were fully grown at 12 years of age, others were still
slowly growing at 20 years, and one individual was still rapidly growing at
18 years. The oldest individual found was 27 years and still growing; most
individuals were between 12 and 20 years old.
However, some individually may well have lived much longer, because the
fossils from Frick and Trossingen are all animals that died in accidents,
and not from old age. Due to the absence of individuals smaller than
4.8 metres (16 ft) long, it is not possible to deduce a complete
ontogenetic series for Plateosaurus or
determine the growth rate of animals under 10 years of age.
Return to the
Old Earth Ministries Online Dinosaur
Curriculum homepage.

Shopping
Bay
State Replicas - None
Black
Hills Institute - None
|

.jpg)

