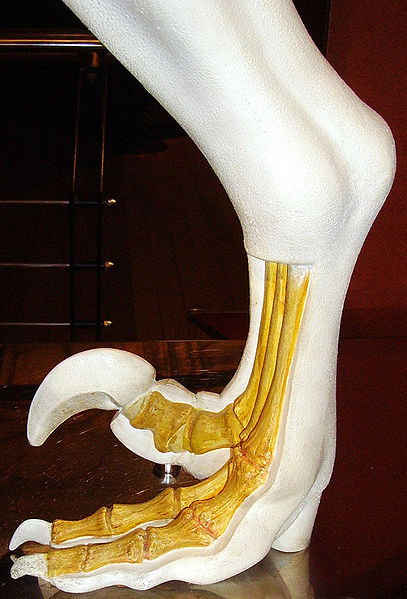
Achillobator).
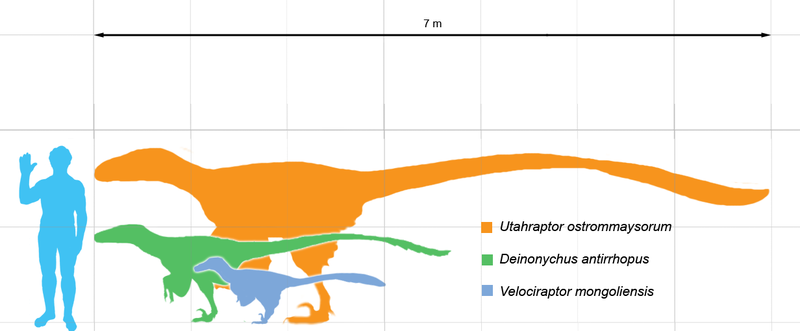
Some may have grown larger; undescribed specimens of
Utahraptor in
BYU collections belonged to individuals that may have reached up to 11 m
(36 ft) long, though these await more detailed study. Large size appears to
have evolved at least twice among dromaeosaurids; once among the
dromaeosaurines Utahraptor and
Achillobator, and again among
the unenlagiines (Austroraptor,
which measured 5 m [16 ft] long). A possible third lineage of giant
dromaeosaurs is represented by isolated teeth found on the
Isle of Wight,
England. The teeth belong
to an animal the size of the dromaeosaurine
Utahraptor, but
they appear to belong to velociraptorines, judging by the shape of the
teeth.
Mahakala is both the most primitive dromaeosaurid ever described
and the smallest. This evidence, combined with the small size of other
primitive relatives such as Microraptor and the
troodontid
Anchiornis, indicates that the common ancestor of dromaeosaurids,
troodontids, and birds – which is called the ancestral
paravian
– may have been very small, at around 65 cm in length and 600 to 700 grams
of mass.
Feathers
There is a large body of evidence showing that dromaeosaurids were
covered in
feathers. Some dromaeosaurid fossils preserve long, pennaceous feathers
on the hands and arms (remiges) and tail (rectrices), as well
as shorter, down-like feathers covering the body. Other fossils, which do
not preserve actual impressions of feathers, still preserve the associated
bumps on the forearm bones where long wing feathers would have attached in
life. Overall, this feather pattern looks very much like
Archaeopteryx.
The first known dromaeosaur with definitive evidence of
feathers
was
Sinornithosaurus, reported from
China by Xu
et al.
in 1999. Many other dromaeosaurid fossils have been found with feathers
covering their bodies, some with fully-developed feathered wings.
Microraptor even
shows evidence of a second pair of wings on the hind legs. While direct
feather impressions are only possible in fine-grained sediments, some
fossils found in coarser rocks show evidence of feathers by the presence of
quill knobs, the attachment points for wing feathers possessed by some
birds. The dromaeosaurids Rahonavis and
Velociraptor have both
been found with quill knobs, showing that these forms had feathers despite
no impressions having been found. In light of this, it is most likely that
even the larger ground-dwelling dromaeosaurids bore feathers, since even
flightless birds today retain most of their plumage, and relatively large
dromaeosaurids, like Velociraptor, are known to have retained
pennaceous feathers. Though some scientists had suggested that the larger
dromaeosaurids lost some or all of their insulatory covering, the discovery
of feathers in Velociraptor specimens has been cited as evidence that
all members of the family retained feathers.
In Popular Culture
Raptors have captured the imagination
of people everywhere. They were a central focus of the Jurassic Park
movie trilogy, and have been portrayed in numerous other movies and
television shows. One novel,
Raptor Red,
by famed paleontologist Bob Bakker, portrays a year in the life of a female
Utahraptor. It is available from
Amazon.
--------------Optional Reading Below this
Point---------------
Paleobiology
Claw function
There is currently disagreement about the function of the enlarged
"sickle claw" on the second toe. When John Ostrom described it for
Deinonychus in 1969, he interpreted the claw as a blade-like slashing
weapon, much like the canines of some
saber-toothed cats, used with powerful kicks to cut into prey. Adams
(1987) suggested that the talon was used to disembowel large
ceratopsian dinosaurs. The interpretation of the sickle claw as a
killing weapon applied to all dromaeosaurids. However, Manning
et al.
argued that the claw instead served as a hook, reconstructing the keratinous
sheath with an elliptical cross section, instead of the previously inferred
inverted teardrop shape. In Manning's interpretation, the second toe claw
would be used as a climbing aid when subduing bigger prey and also as
stabbing weapon.
Ostrom compared
Deinonychus to the
ostrich and
cassowary. He noted
that the bird species can inflict serious injury with the large claw on the
second toe. The
cassowary
has claws up to 125 millimetres (4.9 in) long. Ostrom cited Gilliard (1958)
in saying that they can sever an arm or disembowel a man. Kofron (1999 and
2003) studied 241 documented cassowary attacks and found that one human and
two dogs had been killed, but no evidence that
cassowaries can disembowel or dismember other animals.
Cassowaries use their claws to defend themselves, to attack threatening
animals, and in agonistic displays such as the Bowed Threat Display. The
seriema also has an
enlarged second toe claw, and uses it to tear apart small prey items for
swallowing.
Phillip Manning and colleagues (2009) attempted to test the function of
the sickle claw and similarly shaped claws on the forelimbs. They analyzed
the bio-mechanics of how stresses and strains would be distributed along the
claws and into the limbs, using X-ray imaging to create a three dimensional
contour map of a forelimb claw from
Velociraptor.
For comparison, they analyzed the construction of a claw from a modern
predatory bird, the
Eagle Owl. They found that, based on the way that stress was conducted
along the claw, they were ideal for climbing. The scientists found that the
sharpened tip of the claw was a puncturing and gripping instrument, while
the curved and expanded claw base helped transfer stress loads evenly.
The Manning team also compared the curvature of the dromaeosarid "sickle
claw" on the foot with curvature in modern birds and mammals. Previous
studies had shown that the amount of curvature in a claw corresponded to
what lifestyle the animal has: animals with strongly curved claws of a
certain shape tend to be climbers, while straighter claws indicate
ground-dwelling lifestyles. The sickle-claws of the dromaeosaurid
Deinonychus have a curvature of 160 degrees, well within the range of
climbing animals. The forelimb claws they studied also fell within the
climbing range of curvature.
Paleontologist Peter Mackovicky commented on the Manning team's study,
stating that small, primitive dromaeosaurids (such as
Microraptor) were
likely to have been tree-climbers, but that climbing did not explain why
later, gigantic dromaeosaurids such as
Achillobator
retained highly curved claws when they were too large to have climbed trees.
Mackovickey speculated that giant dromaeosaurids may have adapted the claw
to be used exclusively for latching on to prey.
Group behavior
Deinonychus
fossils have been uncovered in small groups near the remains of the
herbivore
Tenontosaurus, a larger
ornithischian dinosaur. This had been interpreted as evidence that these
dromaeosaurs hunted in coordinated packs like some modern
mammals.
However, not all
paleontologists found the evidence conclusive, and a subsequent study
published in 2007 by Roach and Brinkman suggests that the
Deinonychus
may have actually displayed a disorganized mobbing behavior. Modern
diapsids,
including
birds
and
crocodiles (the closest relatives of dromaeosaurs), display minimal
cooperative hunting; instead, they are usually either solitary hunters, or
are drawn to previously-killed carcasses, where conflict often occurs
between individuals of the same species. For example, in situations where
groups of
komodo dragons are eating together, the largest individuals eat first
and might attack smaller komodo dragons that attempt to feed; if the smaller
animal dies, it is usually
cannibalized. When this information is applied to the sites containing
putative pack-hunting behavior in dromaeosaurs, it appears somewhat
consistent with a komodo- or crocodile-like feeding strategy.
Deinonychus
skeletal remains found at these sites are from subadults, with missing parts
that may have been eaten by other Deinonychus, which a study by Roach
et al. presented as evidence against the idea that the animals
cooperated in the hunt.
In 2007, scientists described the first known extensive dromaeosaur
trackway, in Shandong, China. In addition to confirming the hypothesis
that the sickle-claw was held retracted off the ground, the trackway (made
by a large,
Achillobator-sized species) showed evidence of six individuals of
about equal size moving together along a shoreline. The individuals were
spaced about one meter apart, and retained the same direction of travel,
walking at a fairly slow pace. The authors of the paper describing these
footprints interpreted the trackways as evidence that some species of
dromaeosaurs lived in groups. While the trackways clearly do not represent
hunting behavior, the idea that groups of dromaeosaurs may have hunted
together could not be ruled out.
Flying and gliding
The ability to
fly or glide has been suggested for at least two dromaeosaurid species.
The first,
Rahonavis
ostromi (originally classified as avian bird, but found to be a
dromaeosaurid in later studies) may have been capable of powered
flight, as indicated by
its long forelimbs with evidence of quill knob attachments for long sturdy
flight feathers. The forelimbs of
Rahonavis were more powerfully
built than Archaeopteryx, and show evidence that they bore strong
ligament attachments necessary for flapping flight. Luis Chiappe concluded
that, given these adaptations, Rahonavis could probably fly but would
have been more clumsy in the air than modern birds.
Another species of dromaeosaurid,
Microraptor gui, may have been capable of gliding using its
well-developed wings on both the fore and hind limbs. A 2005 study by
Sankar Chatterjee
suggested that the wings of Microraptor functioned like a split-level
"biplane", and that it
likely employed a
phugoid
style of gliding, in which it would launch from a perch and swoop downward
in a 'U' shaped curve, then lift again to land on another tree, with the
tail and hind wings helping to control its position and speed. Chatterjee
also found that Microraptor had the basic requirements to sustain
level powered flight in addition to gliding.
Powered flight has also been suggested for the species
Cryptovolans pauli (the name of which means "hidden flyer"), though
Cryptovolans is probably synonymous with
Microraptor.
Classification
Relationship with
birds
Dromaeosaurids share many features with early
birds (clade
Avialae or
Aves). The precise nature of their relationship to birds has undergone a
great deal of study, and hypotheses about that relationship have changed as
large amounts of new evidence became available. As late as 2001, Mark Norell
and colleagues analyzed a large survey of
coelurosaur fossils and produced the tentative result that
dromaeosaurids were most closely related to birds, with
troodontids as a more distant outgroup. They even suggested that
Dromaeosauridae could be
paraphyletic relative to Avialae. In 2002, Hwang and colleagues utilized
the work of Norell et al., including new characters and better fossil
evidence, to determine that birds (avialans) were better thought of as
cousins to the dromaeosaurids and
troodontids.
The current consensus among paleontologists agrees with the findings of
Hwang et al. (2002); that dromaeosaurids are most closely related to
the troodontids, and together with the troodontids form the clade
Deinonychosauria.
Deinonychosaurians in turn are the
sister taxon to avialans, and therefore the closest relatives of avialan
birds. A consensus of paleontologists has concluded that there is not yet
enough evidence to determine whether any dromaeosaurs could fly or glide, or
whether they evolved from ancestors that could.
Alternative Theories and Flightlessness
Dromaeosaurids are so birdlike that they have led some researchers to
argue that they would be better classified as birds. First, since they had
feathers, dromaeosaurs (along with many other coelurosaurian
theropod dinosaurs) are “birds” under traditional definitions of the
word “bird”, or “Aves”, that are based on the possession of feathers.
However, other scientists, such as Lawrence Witmer, have argued that calling
a theropod like
Caudipteryx a bird because it has feathers may stretch the word past
any useful meaning.
At least two schools of researchers have proposed that dromaeosaurs may
actually be descended from flying ancestors. Hypotheses involving a flying
ancestor for dromaeosaurs are sometimes called “Birds
Came First” (BCF).
George Olshevsky
is usually credited as the first author of BCF. In his own work, Gregory S.
Paul pointed out numerous features of the dromaeosaurid skeleton that he
interpreted as evidence that the entire group had evolved from flying,
dinosaurian, ancestors, perhaps something like
Archaeopteryx.
In that case, the larger dromaeosaurids were secondarily flightless, like
the modern
ostrich. In
1988, Paul suggested that dromaeosaurids may actually be more closely
related to modern birds than to Archaeopteryx. By 2002, however, Paul
placed dromaeosaurs and Archaeopteryx as the closest relatives to one
another.
In 2002, Hwang
et al. found that
Microraptor was the most
primitive dromaeosaur. Xu and colleagues in 2003 cited the basal position of
Microraptor, along with feather and wing features, as evidence that
the ancestral dromaeosaur could glide. In that case the larger dromaeosaurs
would be secondarily terrestrial—having lost the ability to glide later in
their evolutionary history.
Also in 2002, Steven Czerkas described
Cryptovolans,
though it is a probable junior synonym of
Microraptor. He
reconstructed the fossil inaccurately with only two wings and thus argued
that dromaeosaurs were proper birds, rather than possible gliders. He later
issued a revised reconstruction in agreement with that of
Microraptor
Other researchers, like
Larry Martin believe
that dromaeosaurs, along with all maniraptorans are not dinosaurs at all.
Martin asserted for decades that birds were unrelated to maniraptorans, but
in 2004 he changed his position, and now he agrees that the two are the
closest of relatives. Martin believes that maniraptorans are secondarily
flightless birds, and that birds evolved from non–dinosaurian archosaurs, so
that most of the species formerly called theropods would now not even be
classified as dinosaurs.
In 2005, Mayr and Peters described the anatomy of a very well preserved
specimen of Archaeopteryx, and determined that its anatomy was more
like non-avian theropods than previously understood. Specifically, they
found that Archaeopteryx had a primitive palatine, unreversed hallux,
and hyper-extendable second toe. Their phylogenetic analysis produced the
controversial result that
Confuciusornis
was closer to Microraptor than to
Archaeopteryx, making the
Avialae a paraphyletic taxon. They also suggested that the ancestral
paravian was able to fly or glide, and that the dromaeosaurs and troodontids
were secondarily flightless (or had lost the ability to glide). Corfe and
Butler criticized this work on methodological grounds.
A challenge to all of these alternative scenarios came when Turner and
colleagues in 2007 described a new dromaeosaurid,
Mahakala, which they found to be the most basal and most primitive
member of the Dromaeosauridae, more primitive than
Microraptor.
Mahakala had short arms and no ability to glide. Turner
et al.
also inferred that flight evolved only in the Avialae, and these two points
suggested that the ancestral dromaeosaurid could not glide or fly. Based on
this cladistic analysis, Mahakala suggests that the ancestral
condition for dromaeosaurids is non-volant.
Return to the
Old Earth Ministries Online Dinosaur
Curriculum homepage.