|
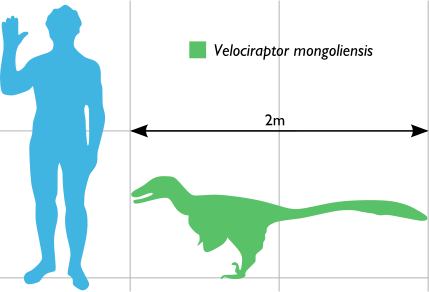
Smaller than other dromaeosaurids
like Deinonychus
and Achillobator,
the turkey-sized Velociraptor
nevertheless shared many of the same anatomical features. It was a bipedal,
feathered carnivore with a long, stiffened tail
and an enlarged sickle-shaped claw on each hindfoot, which is thought to
have been used to kill its prey. Velociraptor
can be distinguished from other dromaeosaurids by its long and low skull,
with an upturned snout.
Velociraptor
(commonly shortened to 'raptor') is one of the dinosaur genera most familiar to the general public due to its prominent role in the
Jurassic Park motion
picture series. In the films it was shown with anatomical inaccuracies,
including being much larger than it was in reality and without feathers. It
is also well known to paleontologists, with over a dozen described fossil
skeletons—the most of any dromaeosaurid. One particularly famous specimen
preserves a Velociraptor
locked in combat with a
Protoceratops.
Velociraptor
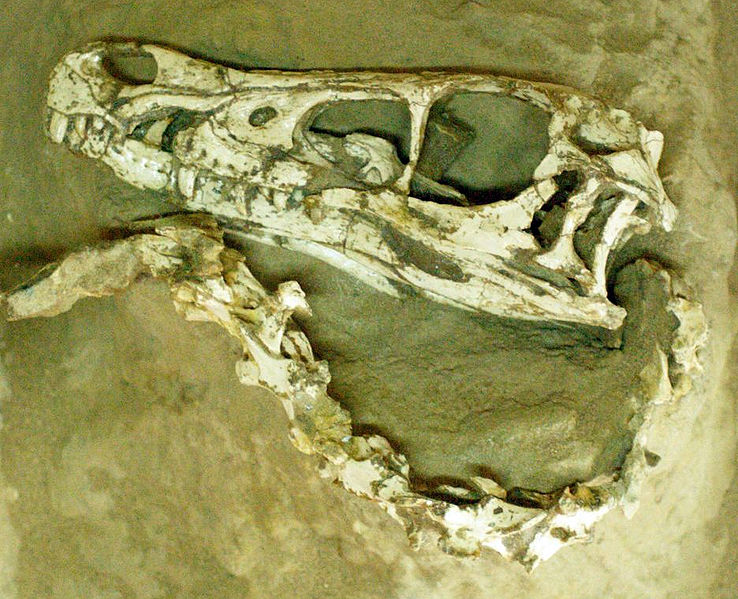
was a mid-sized dromaeosaurid, with adults measuring up to 2.07 m (6.8 ft) long, 0.5 m (1.6 ft) high at the hip, and weighing up to 15 kg (33 lb). The
skull, which grew up to 25 cm (9.8 in) long, was uniquely up-curved, concave
on the upper surface and convex on the lower. The jaws were lined with 26–28
widely spaced teeth on each side, each more strongly serrated on the back
edge than the front—possibly an adaptation that improved its ability to
catch and hold fast-moving prey.
Velociraptor,
like other dromaeosaurids, had a large manus ('hand') with three strongly
curved claws, which were similar in construction and flexibility to the wing
bones of modern birds. The second digit was the longest of the three digits
present, while the first was shortest. The structure of the carpal (wrist)
bones prevented
pronation of the wrist
and forced the 'hands' to be held with the
palmar
surface facing inwards (medially),
not downwards. The first digit of the foot, as in other theropods, was a
small
dewclaw. However,
whereas most theropods had feet with three digits contacting the ground,
dromaeosaurids like Velociraptor
walked on only their third and fourth digits. The second digit, for which
Velociraptor is
most famous, was highly modified and held retracted off of the ground. It
bore a relatively large, sickle-shaped claw, typical of dromaeosaurid and
troodontid dinosaurs. This enlarged claw, which could be over 6.5 cm (2.6
in) long around its outer edge, was most likely a predatory device used to
tear into prey, possibly delivering a fatal blow.
Long bony projections
(prezygapophyses) on the upper surfaces of the vertebrae, as well as
ossified
tendons underneath,
stiffened the tail of Velociraptor.
The prezygapophyses began on the tenth tail (caudal) vertebra and extended
forward to brace four to ten additional vertebrae, depending on position in
the tail. The stiffening forced the entire tail to act as a single rod-like
unit, preventing vertical motion between vertebrae. However, at least one
specimen preserves a series of intact tail vertebrae curved sideways into an
S-shape, suggesting
that there was considerably more horizontal flexibility. These adaptations
of the tail probably provided balance and stability while turning,
especially at high speeds.
In 2007, paleontologists reported
the discovery of quill knobs on a well-preserved
Velociraptor mongoliensis
forearm from Mongolia, confirming the presence of
feathers in this
species.
History
During an
American Museum of Natural History
expedition to the Outer Mongolian
Gobi
Desert, on 11 August 1923 Peter Kaisen
recovered the first Velociraptor
fossil known to science: a crushed but complete skull, associated with one
of the raptorial second toe claws (AMNH
6515). In 1924, museum president
Henry Fairfield Osborn
designated the skull and claw (which he assumed to come from the hand) as
the type specimen of his
new genus,
Velociraptor.
This name is derived from the
Latin
words velox
('swift') and raptor
('robber' or 'plunderer') and refers to the animal's
cursorial nature and carnivorous diet.
Osborn named the type species V. mongoliensis
after its country of origin. Earlier that year, Osborn had mentioned the
animal in a popular press article, under the name "Ovoraptor djadochtari"
(not to be confused with the similarly named
Oviraptor). However, because the name
"Ovoraptor" was not published in a scientific journal or accompanied by a
formal description, it is considered a
nomen nudum ('naked name'), and the name
Velociraptor
retains priority.
While North American teams were shut
out of communist Mongolia
during the Cold War, expeditions
by Soviet and Polish scientists, in
collaboration with Mongolian colleagues, recovered several more specimens of
Velociraptor. The
most famous is part of the legendary "Fighting Dinosaurs" specimen (GIN
100/25), discovered by a Polish-Mongolian team in 1971. This fossil
preserves a single Velociraptor
in the midst of battle against a lone
Protoceratops. This specimen is considered a
national treasure of Mongolia, although in 2000 it was loaned to the
American Museum of Natural History in
New York City for a temporary exhibition.
Between 1988 and 1990, a joint
Chinese-Canadian
team discovered Velociraptor
remains in northern China. American scientists returned to Mongolia in 1990,
and a joint Mongolian-American expedition to the Gobi, led by the American
Museum of Natural History and the Mongolian Academy of Sciences,
turned up several well-preserved skeletons.
Provenance
All known specimens of
Velociraptor mongoliensis
were discovered in the
Djadochta Formation
(also spelled Djadokhta), in the Mongolian province of Ömnögovi. Species of
Velociraptor, have
also been reported from the slightly younger
Barun Goyot Formation
of Mongolia, though these are indeterminate and may belong to a related
genus instead. These geologic formations are
estimated to date back to the
Campanian
stage (between 83 and
70 million years ago) of the
Late Cretaceous
Epoch.
Paleoecology
All of the fossil sites that have
yielded Velociraptor
remains preserve an arid environment with fields of sand dunes and only
intermittent streams, although the
younger Barun Goyot environment seems to have been slightly wetter than the
older Djadochta. The posture of some complete fossils, as well as the mode
of preservation most show within structureless
sandstone deposits, may show that a number
of specimens were buried alive during sandstorm events common
to the three environments.
Classification
Velociraptor
is a member of the subfamily Velociraptorinae, a
derived sub-group of
the larger family Dromaeosauridae. In
phylogenetic taxonomy,
Velociraptorinae is usually defined as "all dromaeosaurs more closely
related to Velociraptor
than to
Dromaeosaurus."
Dromaeosaurid classification is highly variable. Originally, the subfamily
Velociraptorinae was erected solely to contain
Velociraptor. Other analyses have included other
genera, usually Deinonychus
and
Saurornitholestes. A
recent
cladistic analysis
indicated a
monophyletic
Velociraptorinae containing Velociraptor,
Deinonychus,
Tsaagan, and a closely
related (but uncertainly positioned)
Saurornitholestes.
In the past, other dromaeosaurid
species, including Deinonychus antirrhopus
and Saurornitholestes langstoni,
have sometimes been classified in the genus
Velociraptor. Since
Velociraptor was the first
to be named, these species were renamed
Velociraptor antirrhopus and
V. langstoni. However, the
only currently recognized species of
Velociraptor are V.
mongoliensis and V.
osmolskae.
When first described in 1924,
Velociraptor was placed in
the family
Megalosauridae, as was
the case with most carnivorous dinosaurs at the time (Megalosauridae, like
Megalosaurus,
functioned as a sort of 'wastebin' taxon, where many unrelated species were
grouped together). As dinosaur discoveries multiplied,
Velociraptor was later
recognized as a dromaeosaurid. All dromaeosaurids have also been referred to
the family
Archaeopterygidae by at
least one author (which would, in effect, make
Velociraptor a flightless bird).
Paleobiology
Predatory
behavior
The "Fighting Dinosaurs" specimen,
found in 1971, preserves a Velociraptor
mongoliensis
and
Protoceratops andrewsi
in combat and provides direct evidence of predatory behavior. When
originally reported, it was hypothesized that the two animals drowned.
However, as the animals were preserved in ancient sand dune deposits, it is
now thought that the animals were buried in sand, either from a collapsing
dune or in a sandstorm. Burial must
have been extremely fast, judging from the lifelike poses in which the
animals were preserved. Both forelimbs and one hindlimb of the
Protoceratops are missing,
which has been seen as evidence of scavenging by other
animals.
The distinctive claw, on the second
digit of dromaeosaurids, has traditionally been depicted as a slashing
weapon; its assumed use being to cut and disembowel prey. In the
"Fighting Dinosaurs" specimen, the
Velociraptor
lies underneath, with one of its sickle claws apparently embedded in the
throat of its prey, while the beak of
Protoceratops is clamped down upon the right
forelimb of its attacker. This suggests
Velociraptor may have used its sickle claw to
pierce vital organs of the throat, such as the
jugular vein,
carotid artery, or trachea (windpipe),
rather than slashing the abdomen. The inside edge of the claw was rounded
and not unusually sharp, which may have precluded any sort of cutting or
slashing action, although only the bony core of the claw is known. The thick
abdominal wall of skin and muscle
of large prey species would have been difficult to slash without a
specialized cutting surface. The slashing
hypothesis was tested during a 2005 BBC
documentary,
The Truth About Killer Dinosaurs.
The producers of the program created an artificial
Velociraptor leg with a
sickle claw and used a pork
belly to simulate the dinosaur's prey.
Though the sickle claw did penetrate the abdominal wall, it was unable to
tear it open, indicating that the claw was not used to disembowel prey.
However, this experiment has not been published or repeated by other
scientists, so its results cannot be confirmed.
Remains of
Deinonychus, a closely related
dromaeosaurid, have commonly been found in aggregations of several
individuals. Deinonychus
has also been found in association with a large herbivore,
Tenontosaurus, which has been seen as
evidence of cooperative hunting. The only solid evidence for social behavior
among dromaeosaurids comes from a Chinese trackway of fossil footprints,
which shows six individuals of a large species moving as a group, though no
evidence of cooperative hunting was found. Although many isolated fossils of
Velociraptor have
been found in Mongolia, none were closely associated with any other
individuals. Therefore, while Velociraptor
is commonly depicted as a pack
hunter, as in
Jurassic Park, there is only limited fossil
evidence to support this theory for dromaeosaurids in general, and none
specific to Velociraptor
itself. The pack hunting theory was based on a discovery of several
specimens of
Deinonychus found
around the remains of a
Tenontosaurus. No other group of
dromaeosaurids has been found in close association.
Scavenging
behavior
In 2010 Hone and colleagues
published a paper on their 2008 discovery of shed teeth of what they
believed to be a Velociraptor
near a tooth-marked jaw bone of what they believed to be a
Protoceratops in the Bayan
Mandahu Formation. The authors concluded that the find represented
"late-stage carcass consumption by Velociraptor"
as the predator would have eaten other parts of a freshly-killed
Protoceratops before
biting in the jaw area. The evidence was seen as supporting the inference
from the "Fighting Dinosaurs" fossil that
Protoceratops was part of the diet of
Velociraptor.
Metabolism
Velociraptor
was probably warm-blooded to some
degree, as it required a significant amount of energy to hunt. Modern
animals that possess feathery or furry coats, like
Velociraptor did, tend to
be warm-blooded, since these coverings function as insulation. However, bone
growth rates in dromaeosaurids and some early birds suggest a more moderate
metabolism, compared
with most modern warm-blooded mammals and birds. The
kiwi
is similar to dromaeosaurids in anatomy, feather type, bone structure and
even the narrow anatomy of the nasal passages (usually a key indicator of
metabolism). The kiwi is a highly active, if specialized, flightless bird,
with a stable body temperature and a fairly low resting metabolic rate,
making it a good model for the metabolism of primitive birds and
dromaeosaurids.
Feathers
Fossils of dromaeosaurids more
primitive than Velociraptor
are known to have had feathers covering their
bodies, and fully developed, feathered wings. The fact that the ancestors of
Velociraptor were
feathered and possibly capable of flight long suggested to paleontologists
that Velociraptor
bore feathers as well, since even flightless birds today retain most of
their feathers.
In September 2007, researchers found
quill knobs on the
forearm of a Velociraptor
found in Mongolia. These bumps on bird wing bones show where feathers
anchor, and their presence on Velociraptor
indicate it too had feathers. The
forearm specimen on which the quill knobs were found (specimen number IGM
100/981) represents an animal 1.5 meters in length (5 ft) and 15 kilograms
(33 lbs) in weight. Based on the spacing of the six preserved knobs in this
specimen, the authors suggested that
Velociraptor bore 14 secondaries (wing feathers
stemming from the forearm), compared with 12 or more in
Archaeopteryx, 18 in
Microraptor, and 10 in
Rahonavis. This type of variation in the
number of wing feathers between closely related species, the authors
asserted, is to be expected, given similar variation among modern birds.
Velociraptor
is presumed to have been flightless due to its relatively large size and short
forelimbs, yet the evidence implies that the ancestors of dromaeosaurids could fly,
making Velociraptor
and other large members of this family secondarily flightless, though it is
possible the large wing feathers inferred in the ancestors of
Velociraptor had a purpose
other than flight. The feathers of the flightless
Velociraptor may have been
used for display, for covering their nests while brooding, or for added
speed and thrust when running up inclined slopes.
In popular
culture
Velociraptor
are well-known for their role as vicious and cunning killers in the 1990
novel
Jurassic Park by
Michael Crichton and
its 1993 film adaptation,
directed by Steven Spielberg, in
which they served as the main antagonists. The "raptors" portrayed in
Jurassic Park were modeled
after a larger relative,
Deinonychus, which Gregory Paul at the
time called
Velociraptor antirrhopus.
The paleontologists in the film and the novel excavate a so-called
Velociraptor skeleton in
Montana, far from the central Asian range of
Velociraptor but
well within the range of Deinonychus.
A character in Crichton's novel also states that "…Deinonychus
is now considered one of the velociraptors", indicating that Crichton used
Paul's taxonomy, though the
"raptors" in the novel are referred to as V.
mongoliensis.
The filmmakers also had the size of
the film's Velociraptor
increased for dramatic reasons, and changed the shape of the snout.
Additionally, the forelimbs of the film animals differed in structure and
posture from those of real dromaeosaurids and their tails were too short and
flexible, anatomical errors which directly contradict fossil evidence.
Another noticeable difference is the lack of feathers for the novel and film
version of Velociraptor.
In real life, Velociraptor,
like many other maniraptoran theropods,
was covered in feathers.
Jurassic Park and
The Lost World: Jurassic Park,
however, were released before this was known. In
Jurassic Park III, the
Velociraptor are
depicted with quill-like structures along the back of the head and neck,
although these do not resemble the down-like feathers known from real-life
dromaeosaurids, and the quill knobs on some
Velociraptor specimens show that they had fully
developed feathers akin to those of modern birds. Also in
Jurassic Park III, Dr. Alan Grant, played
by Sam Neill, states that
Velociraptor were
smarter than dolphins, whales and some primates. Based on fossil evidence,
this is highly unlikely.
Due to the success of many
Jurassic Park-related
products, Velociraptor
has become a ubiquitous representation of dinosaurs in popular culture. It
has been featured in numerous
toy lines, animated films, video
games and television series for
children, along with several television documentaries.
In 1995, the city of Toronto was awarded a
National Basketball Association
expansion team, which was named the
Toronto Raptors.
Return to the
Old Earth Ministries Online Dinosaur
Curriculum homepage.

Shopping
Bay
State Replicas - Skulls, Leg, Foot, Claws
Black
Hills Institute - Velociraptor vs. protoceratops fighting
|